
過去普遍認為,脂肪組織僅是身體儲存備用能量的倉庫,然而從內分泌角度來看,脂肪組織不僅負責儲存熱量,同時也是活躍的內分泌器官。它持續分泌多種荷爾蒙與訊號分子,與大腦、腸道、肝臟及腎上腺等器官維持雙向溝通,共同調控食慾、能量代謝與脂肪分布。
然而,在長期肥胖與慢性壓力的環境下,訊號持續分泌,分泌量高於正常值,但大腦卻逐漸失去解讀訊號的能力,是造成許多人明明飲食受控、體重卻難以下降的核心原因之一。
瘦素是什麼?並非「燃脂素」,而是脂肪量訊號
瘦素(leptin)是一種蛋白質荷爾蒙,早期由小鼠的肥胖基因中率先發現,原名意為「纖細」,反映了早期研究者的期待:一種能讓身體自動抵抗肥胖的訊號分子。
瘦素主要由白色脂肪組織(White Adipose Tissue, WAT)的脂肪細胞分泌,其分泌量與脂肪儲備量呈正比,即脂肪細胞越多、脂質含量越高,製造的瘦素也越多。瘦素進入血液循環後必須穿越血腦屏障(BBB)才能到達其主要作用部位:下視丘的弓狀核(Arcuate Nucleus),並與LepRb受體結合,向大腦傳遞身體所儲備能量的多寡,調整食慾行為與代謝輸出(又稱總熱量消耗,簡稱TDEE)。
瘦素的分泌具有明顯的晝夜節律:在夜晚睡眠期間達到高峰,抑制夜間食慾並加速代謝,而白天午後降至最低。此與睡眠品質及進食時段密切相關,因此睡眠型態的紊亂也是干擾瘦素分泌的原因之一。
瘦素機制正常運作下,身體趨向能量平衡:
步驟一、體脂增加,促進瘦素分泌上升,大腦收到「儲備充足」訊號
步驟二、食慾受到抑制、能量消耗微幅提升
步驟三、負回饋影響,導致體脂緩慢回落
然而,若脂肪細胞持續增加,瘦素阻抗一旦形成,此機制便開始失去功能。
下表整理瘦素的基本特性與核心功能:
| 特性 | 說明 |
| 主要來源 | 白色脂肪組織(皮下脂肪為主) |
| 主要作用部位 | 下視丘弓狀核(LepRb 受體),透過JAK2-STAT3路徑傳訊 |
| 核心功能 | 向大腦回報脂肪儲備現況;抑制食慾、調升能量消耗 |
| 分泌節律 | 夜間睡眠期達高峰,午後最低 |
| 肥胖時的表現 | 血清濃度顯著升高,但大腦對訊號的接收與回應能力受損 |
一、愈胖瘦素愈高?瘦素阻抗如何讓大腦收不到飽足訊號?
發表於《New England Journal of Medicine》的人體研究證實,肥胖者的血清瘦素濃度顯著高於正常體重者,且與體脂百分比呈強正相關(r=0.85,p<0.001),而後續多項研究反覆驗證此正相關性:隨著肥胖程度上升,血清瘦素濃度同步增加,意即脂肪細胞越多,分泌的瘦素自然越多。
因此,肥胖問題並不在於瘦素「不夠多」,而是大腦已無法正常接收與回應此訊號,此現象稱為「瘦素阻抗」(Leptin Resistance)。過程與第二型糖尿病中的胰島素阻抗高度相似:荷爾蒙確實有分泌、濃度甚至偏高,但目標細胞對其反應能力已大幅下降。2024 年《Journal of Clinical Investigation》的回顧文章指出,瘦素阻抗幾乎普遍存在於大多數肥胖案例之中,被視為一種系統性的代謝缺陷。
刊登於《PLoS One》的動物實驗顯示,當人工固定肥胖小鼠的血清瘦素在低水平,即使長期餵食高脂飲食,小鼠仍維持對瘦素的高敏感性;而允許瘦素自然上升的組別,則出現顯著阻抗。因此高脂飲食本身,不足以獨立引發阻抗,高濃度瘦素才是關鍵觸發阻抗的條件。
下視丘如何決定要不要停止進食?
瘦素要發揮食慾調節作用,僅試圖「提升瘦素濃度」並不是解方,核心在於恢復大腦對於訊號的敏感度。瘦素需成功進入血腦屏障、抵達對應腦區、結合瘦素受體,並啟動受體後方的細胞訊號路徑。上述任何一環節失效,都會讓「脂肪庫存已充足」的訊息無法完整傳遞至大腦。
| 障礙層次 | 機制說明 | 結果 |
| 血腦屏障阻斷(周邊阻抗) | 肥胖相關高三酸甘油酯血症,抑制瘦素受體轉運效率 | 大量瘦素滯留血液,無法進入腦部 |
| 受體去敏化(中樞阻抗) | 慢性高瘦素導致訊息傳遞路徑過度活化,並產生負回饋,主動封鎖受體傳訊 | 瘦素即使入腦,神經元也難以正常回應 |
| 神經元功能受損(中樞阻抗) | 高脂飲食與慢性發炎誘發下視丘壓力,神經元活性下降 | 飽足訊號強度持續弱化,越吃越多、代謝減緩 |
| 發炎與細胞壓力升高 | 免疫細胞過度活化,釋放大量促發炎細胞激素(如 TNF-α、IL-6) | 進一步干擾瘦素敏感度,形成代謝惡性循環 |
目前臨床上雖尚未建立「瘦素阻抗」的標準診斷指標,但從大多數肥胖者的日常情境來看,難以抑制的飢餓感、對飲食控制反應遲鈍,相當程度可能是瘦素被多層阻斷機制的具體表現,而非單純的行為意志力問題。而壓力荷爾蒙(皮質醇)會更加重此現象。
二、壓力皮質醇如何介入?HPA軸讓食慾、血糖與腹部脂肪一起失衡
面對壓力時,會引起下視丘、腦下垂體與腎上腺(HPA軸)反應,促使腎上腺分泌皮質醇。正常狀態下,皮質醇遵循嚴格的晝夜節律:清醒後30至45分鐘內迅速攀升至全日高點(稱為「皮質醇清醒反應」,Cortisol Awakening Response),隨後在白天穩定下降,至夜間睡眠期降至最低值。「早高晚低」的變化,是協調全身代謝節律的核心訊號,也是維持胰島素敏感性與能量平衡的重要條件之一。
慢性壓力會系統性地破壞這個節律。HPA軸長期活化後,皮質醇分泌型態會出現明顯的「扁平化」:清晨峰值降低或延遲,而夜間本應降至最低的濃度反而異常偏高,晝夜濃度差距縮小,讓身體長時間曝露在相對偏高的皮質醇環境中,導致代謝異常,引發「壓力胖」、肌肉流失、胰島素阻抗及免疫力下降。
皮質醇與腹部脂肪堆積高度相關
內臟脂肪組織中的糖皮質激素受體(GR)密度,顯著高於皮下脂肪。意即同樣濃度的皮質醇,在腹部脂肪中能引發更強烈的反應,促進脂肪細胞分化與脂質儲存。
另一關鍵是11β-HSD1酵素(11β-羥類固醇脫氫酶第一型),負責將非活性的可體松(cortisone)轉化為活性皮質醇(cortisol),其在內臟脂肪中活性特別高。透過局部合成,內臟持續獲得較高濃度的活性皮質醇,不斷刺激脂肪細胞增生,刺激免疫細胞過度活化,釋放大量促發炎細胞激素(如 TNF-α、IL-6),進一步加重肥胖與胰島素阻抗。
瘦素與胰島素都會將身體能量狀態傳給大腦與周邊組織。當兩種訊號都變遲鈍時,就可能同時出現「脂肪量增加、飽足訊號不清楚、血糖調節壓力升高、脂肪儲存傾向增加」的代謝困境。
三、腸道菌相失衡加重代謝危機:腸漏、發炎、胰島素阻抗讓飽足訊號更不敏感
健康的腸道屏障由一層緊密連結的腸上皮細胞組成,選擇性允許營養素通過,同時阻止細菌毒素與代謝廢物滲入體循環。腸道菌相失衡會損傷緊密的連接系統,造成腸道通透性增加,稱為「腸漏」(Leaky Gut)。
腸漏形成後,革蘭氏陰性菌外膜的主要毒素成分脂多醣(lipopolysaccharide, LPS)得以進入血液循環,與內臟脂肪直接釋放的促發炎訊號形成疊加效應,加劇發炎,持續損害胰島素與瘦素的訊號傳遞效率。
多因子交互作用,形成代謝惡性循環
- 慢性低度發炎:干擾脂肪組織、下視丘與受體後訊號傳遞。導致瘦素訊號變遲鈍、食慾調節困難、代謝壓力升高。
- 胰島素阻抗:使血糖與脂肪儲存調節變得更費力。
- 腸道菌相失衡:導致發炎反應、胰島素敏感度下降、能量代謝失衡。
- 中樞(下視丘)發炎:影響大腦對瘦素與營養訊號的反應。飽足訊號下降、能量消耗調節受影響。
四、顧好腸道健康:穩定皮質醇節律與代謝平衡
慢性壓力不只透過皮質醇影響脂肪分布,HPA軸的過度活化已被證實會干擾腸道微生態的平衡,增加腸漏,並加重菌相失調的程度。反之,腸道菌相失調引起的系統性發炎,也會增加皮質醇分泌。
因此,腸道菌相的調節不只攸關消化系統本身,更是緩解系統性發炎、間接支持皮質醇節律的關鍵之一。
植物乳桿菌K21:調節脂肪代謝、降低瘦素與強化腸道屏障
在特定益生菌對於代謝調節的研究領域中,具有潛在作用的植物乳桿菌K21(Lactobacillus plantarum K21)菌株,展現相當優異的實驗結果。研究以高脂飲食誘發肥胖(DIO)的雄性小鼠為模型,分為正常飲食(ND)、高脂飲食(HFD),以及高脂飲食加每日灌餵 K21(10⁹ CFU)持續八週。主要研究結果如下:
| 觀察指標 | HFD 組 | HFD + K21 組 |
| 體重增加幅度 | 顯著高於正常飲食 | 增幅受到抑制 |
| 副睪周圍脂肪量 | 大量累積 | 累積量降低 |
| 血漿瘦素濃度 | 顯著升高 | 降低 |
| 膽固醇與三酸甘油酯 | 升高 | 降低 |
| 肝損傷指標(AST/ALT) | 升高,肝損傷明顯 | 改善,肝損傷緩解 |
| 肝臟 PPAR-γ 表達 | 升高 | 顯著下調 |
| 腸道通透性 | 受損 | 強化,屏障功能改善 |
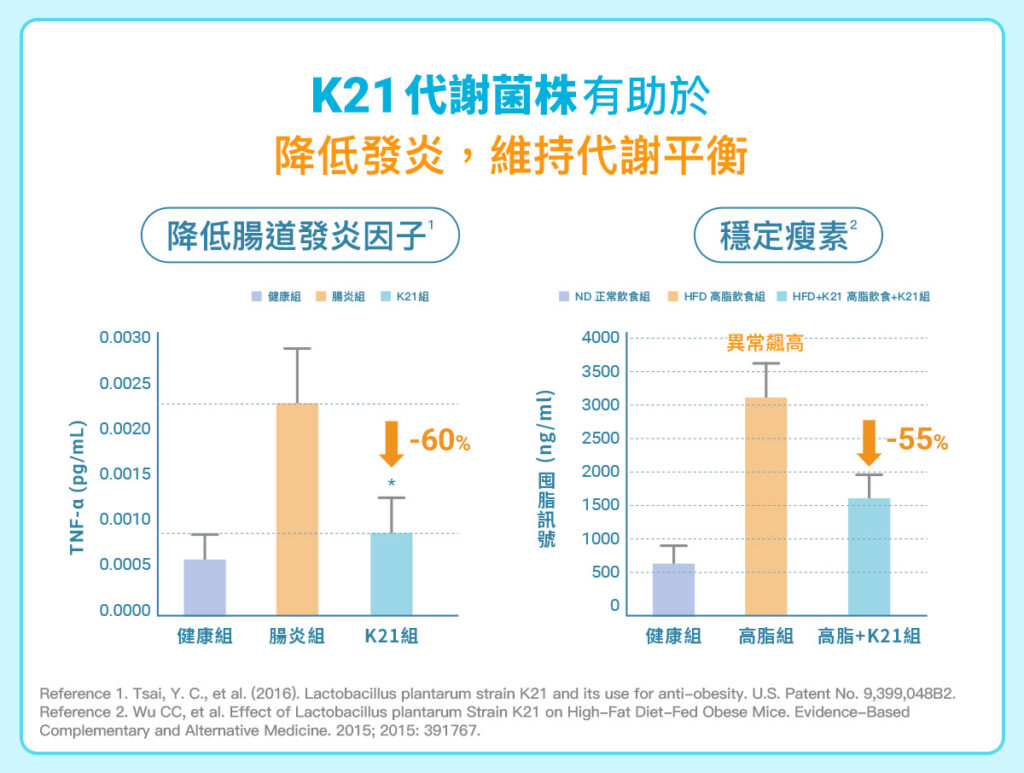
K21 菌株的介入同時觀察到多個與代謝調節相關的變化,包括降低血漿瘦素濃度、減少體重增加與脂肪累積、降低血中膽固醇與三酸甘油酯,並改善腸道屏障相關指標。其中,高三酸甘油酯可能干擾瘦素通過血腦屏障,進而影響大腦接收瘦素訊號的效率。
K21菌株並非單純「抑制肥胖」,而是透過血脂代謝、脂肪堆積、瘦素訊號與腸道屏障等多個環節,共同降低高脂飲食下的代謝壓力。當腸道屏障較完整,細菌的發炎刺激物較不易進入血液;當血脂與脂肪累積壓力下降,瘦素與胰島素訊號也較不易被干擾。
對於瘦素阻抗、腹部脂肪堆積與慢性低度發炎交織而成的代謝危機,理想的策略不應只是追求單一指標的降低,而是回到「降低代謝壓力、維持腸道屏障、支持訊號敏感度」的源頭,進行系統性的整體調節。
參考資料
- Adam, E. K., Quinn, M. E., Tavernier, R., McQuillan, M. T., Dawood, K., & Gilbert, K. E. (2017). Diurnal cortisol slopes and mental and physical health outcomes: A systematic review and meta-analysis. Psychoneuroendocrinology, 83, 25–41.
- Alzamil, H. (2020). Elevated serum TNF-α is related to obesity in type 2 diabetes mellitus and is associated with glycemic control and insulin resistance. Journal of Diabetes Research, 2020, Article 5076858.
- Cao N, Zhao F, Kwok LY, Wang H, Sun Z. Impact of probiotics on weight loss, glucose and lipid metabolism in overweight or obese women: A meta-analysis of randomized controlled trials. Curr Res Food Sci. 2024 Jul 14;9:100810.
- Banks, W. A., Coon, A. B., Robinson, S. M., Moinuddin, A., Shultz, J. M., Nakaoke, R., & Morley, J. E. (2004). Triglycerides induce leptin resistance at the blood-brain barrier. Diabetes, 53(5), 1253–1260.
- van Loenen MR, Geenen B, Arnoldussen IAC, Kiliaan AJ. Ghrelin as a prominent endocrine factor in stress-induced obesity. Nutr Neurosci. 2022 Jul;25(7):1413-1424.
- Considine, R. V., Sinha, M. K., Heiman, M. L., Kriauciunas, A., Stephens, T. W., Nyce, M. R., Ohannesian, J. P., Marco, C. C., McKee, L. J., Bauer, T. L., & Caro, J. F. (1996). Serum immunoreactive-leptin concentrations in normal-weight and obese humans. New England Journal of Medicine, 334(5), 292–295.
- Flier, J. S., & Maratos-Flier, E. (2024). Leptin physiology and pathophysiology: Knowns and unknowns 30 years after its discovery. Journal of Clinical Investigation, 134(2), e174595.
- Goldsmith, C. C., & Dodd, G. T. (2024). TET2: The fat controller of leptin. Life Metabolism, 3(3), loae019.
- Ķimse, L., Reinis, A., Miķelsone-Jansone, L., Gintere, S., & Krūmiņa, A. (2024). A narrative review of psychobiotics: Probiotics that influence the gut–brain axis. Medicina, 60(4), Article 601.
- Kuckuck, S., van Baak, M. A., & Camps, S. G. J. A. (2023). Glucocorticoids, stress and eating: The mediating role of appetite-regulating hormones. Obesity Reviews, 24 (3), e13539.
- Lengton, R., et al. (2025). Glucocorticoids and HPA axis regulation in the stress–obesity connection: A comprehensive overview of biological, physiological and behavioural dimensions. Clinical Obesity, e12725.
- Melo, H. M., et al. (2017). Triglycerides cross the blood–brain barrier and induce central leptin and insulin receptor resistance. International Journal of Obesity, 41, 1525–1534.
- Mishra, A. K., et al. (2023). Recent insights of obesity-induced gut and adipose tissue dysbiosis in type 2 diabetes. Frontiers in Molecular Biosciences, 10, Article 1224982.
- Morton, G. J., Kaiyala, K. J., Fisher, J. D., Ogimoto, K., Schwartz, M. W., & Wisse, B. E. (2010). Hyperleptinemia is required for the development of leptin resistance. PLoS ONE, 5(6), e11376.
- Ołoń E, Gurgul A, Szmatoła T, Jasielczuk I, Kucharski M, Zubel-Łojek J and Zieba DA (2025) Impact of modulating leptin sensitivity on the transcriptomic profile of adult-derived hypothalamic mouse neurons. Front. Mol. Neurosci. 17:1518737.
- Al-Juhani A, Desoky MS, Almuhaimid AA, Zaheer M, Alhaqbani HF, Abalkhail EA, Alanazi SA, Alzahrani RS, Alrefaai M, Desoky R. Efficacy of Gut Microbiome-Targeted Therapies in Modulating Systemic Inflammation and Low-Grade Chronic Inflammatory States in Adults With Metabolic Disorders: A Systematic Review. Cureus. 2025 Sep 21;17(9):e92881.
- Prajna, R. H., Shivananda Nayak, Priya, V., Shruthi Rai, P., Shivaraja Shankara, Y. M., & Prashanthkumar, G. (2025). The role of TNF-Alpha, IL-6, Adiponectin, and Leptin in inflammation and metabolic dysregulation in type 2 diabetes mellitus. Pharmacognosy Journal, 17(6), 699–702.
- Hu W, Zhu H, Gong F. Leptin and leptin resistance in obesity: current evidence, mechanisms and future directions. Endocr Connect. 2025 Sep 29;14(9):e250521.
- Nakandakare-Maia, E.T., Siqueira, J.S., Caccia, C.M. et al. Differential effects of obesogenic diets components on hypothalamic leptin resistance: a narrative review. Nutrire 51, 24 (2026).
- Simon C, Gronfier C, Schlienger JL, Brandenberger G. Circadian and ultradian variations of leptin in normal man under continuous enteral nutrition: relationship to sleep and body temperature. J Clin Endocrinol Metab. 1998 Jun;83(6):1893-9.
- Smith W, Azevedo EP. Hunger Games: A Modern Battle Between Stress and Appetite. J Neurochem. 2025 Feb;169(2):e70006.
- Wang, X., et al. (2024). Short-chain fatty acids: Bridges between diet, gut microbiota, and health. Journal of Gastroenterology and Hepatology, 39(6).
- Obradovic M, Sudar-Milovanovic E, Soskic S, Essack M, Arya S, Stewart AJ, Gojobori T, Isenovic ER. Leptin and Obesity: Role and Clinical Implication. Front Endocrinol (Lausanne). 2021 May 18;12:585887.
- Wilding, J. P. H. (2019). Leptin resistance: Underlying mechanisms and diagnosis. Diabetes, Metabolic Syndrome and Obesity, 12, 1737–1745.
- Wu, C.-C., Weng, W.-L., Lai, W.-L., Tsai, H.-P., Liu, W.-H., Lee, M.-H., & Tsai, Y.-C. (2015). Effect of Lactobacillus plantarum strain K21 on high-fat diet-fed obese mice. Evidence-Based Complementary and Alternative Medicine, 2015, Article 391767.
- Zhang, Y., Proenca, R., Maffei, M., Barone, M., Leopold, L., & Friedman, J. M. (1994). Positional cloning of the mouse obese gene and its human homologue. Nature, 372, 425–432.
延伸閱讀
壓力型肥胖怎麼減?皮質醇(壓力賀爾蒙)失調與腸道菌相失衡對代謝的影響
健檢紅字年年有?從脂肪內分泌到腸道菌:解析代謝症候群的生理成因與逆轉機制
GLP-1 減重機制揭秘:不打針也能瘦?關鍵在於調節「腸道菌相」